©
1996 г.ЭВОЛЮЦИОНИРУЮЩИЕ СИСТЕМЫ
ЛЕВОСТОРОННЕ–АСИММЕТРИЧНЫ?О.В.Трапезов
“
Грешники на страшном судеПроблеме симметрии–асимметрии посвящена поистине необозримая литература. От учебников и научных монографий до произведений, апеллирующих не столько к чертежу и формуле, сколько к художественному образу, и сочетающих в себе научную достоверность с литературной отточенностью.
Вся ошеломляющая пестрота и разнообразие окружающего нас мира подчинены проявлениям симметрии, о чем удачно в свое время высказался Дж. Ньюмен: “Симметрия устанавливает забавное и удивительное сродство между предметами, явлениями и творениями, внешне, казалось бы, ничем не связанными: земным магнетизмом, женской вуалью, поляризованным светом, естественным отбором, теорией групп, инвариантами и преобразованиями, рабочими привычками пчел в улье, строением пространства, рисунками ваз, квантовой механикой, скарабеями, лепестками цветов, интерференционной картиной рентгеновских лучей, делением клеток, равновесными конфигурациями кристаллов, романскими соборами, снежинками, музыкой, теорией относительности...” [1].
В “Кратком Оксфордском словаре” симметрия определяется как
“красота, обусловленная пропорциональностью частей тела или любого целого, равновесием, подобием, гармонией, согласованностью” (сам термин “симметрия” по–гречески означает “соразмерность”, которую древние философы понимали как частный случай гармонии – согласования частей в рамках целого).Симметрия является одной из наиболее фундаментальных и одной из наиболее общих закономерностей мироздания: неживой, живой природы и общества. Ее математическое выражение – теория групп – была признана одним из самых сильных средств познания первоначально в математике, а позднее – в науке и искусстве [30, 49]. Симметрия в рамках общей теории систем (ОТС) предстает как системная категория, обозначающая свойство системы “С” совпадать с самой собой по признакам “П” после изменений “И”.
Симметрия как общенаучное понятие на одном уровне делится на три типа: структурную, геометрическую и динамическую. На следующем уровне каждый тип симметрии включает классическую и неклассическую симметрии, которые в свою очередь имеют разновидности следующего уровня подчинения. Так, неклассическая симметрия структурного типа в числе других содержит три соподчиненных понятия – антисимметрию, цветную симметрию и криптосимметрию. Каждая из них далее выступает в виде простой и кратной симметрии и т.д. На каждой ветви “дерева” данного понятия можно выбрать и родо–видовые отношения (по вертикали), которые подчиняются закону обратного отношения содержания и объема. Так, на ветви структурной симметрии такими отношениями являются симметрия (вообще) ![]() структурнокристаллографическая
структурнокристаллографическая ![]() неклассическая
неклассическая ![]() антисимметрия
антисимметрия ![]() кратная.
кратная.
Категория “симметрия” дополнена ее противоположностью – категорией “асимметрия”, с необходимостью ею предполагаемой и дополняющей ее до гармоничной пары “симметрия – асимметрия”.
В рамках ОТС асимметрия означает несовпадение по признакам “П” системы “С” после изменений “И”. Так как относительно любой совокупности изменений {И} существуют инвариантные признаки, то необходимым дополнением любой асимметрии будет соответствующая ей симметрия.
Принцип симметрии побудил древнегреческого философа Аристотеля, не представлявшего себе Землю иначе как симметричной, предсказать наличие “неизвестной южной земли” (позднее ею оказалась Антарктида) в противовес континентам в северной полусфере.
Сложные понятия в мире физики благодаря симметрии физических законов и физических систем делаются простыми и изящными. Так, квантовая механика, которой приходится пользоваться на микроскопическом уровне, будучи сложнее классической, в значительной мере упрощается при введении принципа симметрии
. Примером может служить то, что частица описывается не заданием ее положения, а заданием волновой функции.Различие между сахарами глюкозой и галактозой (самый тонкий химический анализ не в состоянии отличить одно вещество от другого, поскольку химическая формула их одна и та же: С
6 Н12 O6) определяется пространственной конфигурацией асимметричного углеродного атома. Каждая из двух таких конфигураций является зеркальным отражением другой. Это различие может определить судьбу ребенка с редкой наследственной болезнью – галактоземией. Такие дети не усваивают галактозу, которой так много в молоке. Если они получают пищу, в которой галактоза заменена другим сахаром (например, глюкозой), то они развиваются нормально, если же получают обычную пищу, – становятся слабоумными.Лишь правизна отличает искусственно созданное в лабораторных условиях вещество декстраникотин от левоникотина, который входит в состав любого табака, но если первое соединение нейтрально для организма человека, то по раковым заболеваниям легких уверено лидируют курильщики.
Живой организм не может жить без витамина С – сразу же наступает цинга. Но точно такое же вещество – с одной лишь разницей: молекулы его зеркально отражены – не оказывает на организм вообще никакого влияния.
Из двух антиподов фенилаланина L–изомер вызывает тяжелые психические заболевания, а его D–изомер безвреден.
L–форма сарколизина активна при онкологических заболеваниях, а D– форма – нейтральна.
Гормональное действие L–адреналина гораздо более сильное, чем его D–изомера
.Из двух антиподов хлоромицетина антибиотическими свойствами обладает только левовращающий изомер.
Различная пространственная конфигурация L– и D–аспарагинов, лейцинов и тирозинов влияет на их усвояемость сельскохозяйственными животными: усваиваются только сладкие на вкус D–формы [38, 39].
Еще в 1880–90 гг. немецкий химик Ф. Эрлих находит большие физиологические различия между L– и D–изомерами лейцина фибрина крови [40].
Среди D– и L–изомеров инсектицида гексахлорциклогексана только L–изомер обладает инсектицидной активностью [42].
В блестящих опытах основоположника современной микробиологии Л.Пастера было установлено, что различать левые и правые формы химических веществ способны уже низшие организмы: колония микроорганизмов, помещенных в рацемический раствор виннокаменной кислоты, выедала ее правую форму, и направление плоскости поляризации все больше и больше поворачивалось налево. Впоследствии К.Нейберг и П.Майер подтверждают это на низших грибах: аспергилл в рацемическом растворе цистеина также усваивает только его D–вращающую форму [40]. В своих двух знаменитых лекциях в Химическом обществе Франции, 20 января и 3 февраля 1860 г., Пастер впервые объясняет природу оптической активности молекулярной асимметрией, когда формы молекул лево– и правовращающих изомеров должны относиться друг к другу как зеркальные изображения или как левый и правый винты. Эта замечательная мысль дала направления для дальнейших исследований в создании классической структурной теории. Сам Пастер считал свое открытие молекулярной дисимметрии указывающим прямой путь к постижению “тайны жизни”, то есть ее сущности и происхождения. “Объединив кристаллографию, химию и молекулярную оптику, Пастер с абсолютной точностью пришел к открытию наиболее общих законов, позволяющих выявить самые характерные признаки живой клетки” [7]. По мнению Дагонье, “... из них именно кристаллография решила судьбу его трудов и открытий” [8]. Открытие Пастером нового феномена – диссимметричности пространственной конфигурации молекулы, явилось итогом интеграции идей и методов трех естественных наук – кристаллографии, оптической поляриметрии и химии.
Попытки объяснить природу симметрии – асимметрии
Нарушение симметрии левого и правого занимает умы исследователей уже более 100 лет. О том, что наш мир все–таки несимметричен и четность в нем нарушена, догадывались задолго до фундаментального открытия Ли и Янга. Так, знаменитый Г.Вейль в 1929 г. высказал гипотезу о том, что вращающаяся частица может быть в одной из двух зеркально отраженных форм – обладать левой или правой спиральностью. Вейль не был физиком–ядерщиком и у него не было никаких опытных данных для такой необычной гипотезы. Он просто построил изящную математическую теорию. Но в то время к ней никто не отнесся всерьез, потому что она не согласовывалась
с законом сохранения четности и требовала от природы асимметричности. Вейль не дожил всего два года до того дня, как закон этот был опровергнут и его теория получила титул пророческой. В самом деле, из нее следовало, что у вращающейся частицы должен быть зеркальный двойник, – и его нашли.В западном мире научно–теоретическое понимание симметрии–асимметрии (инь–янский аналог в восточной философской мысли) в значительной мере формируется в трудах гениального Пастера и его предшественников (Р.–Ж. Аюи, О.Лорана и Г.Деляфосса – в области кристаллографии; Ж.–Б. Био и Дж. Гершеля – в области оптической поляриметрии; Э.Шевреля, Й.Я.Берцелиуса, Ж.–Б. Дюма – в области химии). Задумываясь над проблемой происхождения жизни, Пастер открыл, что “симметрия между левым и правым в живой природе нарушена на молекулярном уровне”, а само зарождение биологической изомерии на этом уровне предопределено “диссимметрической совокупностью Вселенной” [9]. В своих поздних изысканиях Пастер высказывает идею об определяющей роли физики в исследовании процесса биопоэза [11, 12, 13, 14]. И вот уже в новейшей физике в свете идей квантовой механики теории слабых взаимодействий “диссимметрические силы” Пастера получают новое истолкование. Так, В.Гейзенберг в размышлениях о единой теории поля говорит: “О том, как после этого возникает нарушение симметрии, мы имели тогда еще менее ясное представление, чем о раздвоении. Но в ходе наших бесед иногда всплывала идея, что мир в целом, т.е. космос, не обязательно должен быть симметричен по отношению
к операциям, при которых природные законы остаются инвариантными, и, следовательно, наблюдаемое у нейтрино уменьшение симметричности можно, вероятно, объяснить несимметрией космоса” [15].Неоднократно пытались объяснить причину асимметрии разными природными факторами – от действия поляризованного солнечного света и грозовых разрядов до несохранения четности при слабых взаимодействиях. Все объяснения имели общий недостаток: они давали слишком малую степень нарушения симметрии.
Мощным стимулом к изучению проблемы симметрии послужило открытие двойной спирали ДНК – носителя информации, записанной на биоорганических макромолекулах.
Проблема симметрии–асимметрии в биологии (иногда ее формулируют, как проблема упорядоченности, регулярности и соразмерности в строении организмов и их развитии) изучается на двух уровнях: на макроуровне (субклеточный, органоидный, организменный) и на микроуровне (молекулярный, биополимерный). При этом выделяются два методологических подхода в изучении названной проблемы: выяснение причинно–следственных взаимосвязей между пространственной конфигурацией биополимеров и их функциональными свойствами; второе – для чего необходимо свойственное всему живому миру соотношение L– и D–биомолекул, т.е. в чем “целесообразность и полезность” для организма той или иной структуры и функции.
То, что в биологических системах встречается лишь один изомер, объяснить несложно. Но почему только L–изомер? Впервые вопрос о биологической целесообразности существующего соотношения стереоизомеров в современном органическом мире был поставлен Пастером: “Почему возникает определенная диссимметрия, а не противоположная, почему только правый сахар... и левые белки” [18]. Ставя так проблему Пастер был глубоко убежден, что изучение этого вопроса – один из важнейших путей к познанию сущности жизни.
В исследовании путей происхождения современного соотношения L– и D–биомолекул выделяются два направления.
1. Существующее соотношение результат случайного, спонтанно происшедшего события, которое могло разыгрываться по двум схемам:
а) возникшая киральная протоструктура уже представляла собой готовую самореплицирующуюся молекулярную систему, содержавшую чистые L–аминокислоты и D–пентозу;
б) в результате стохастического нарушения рацемического баланса одни из изомеров получили перевес над другими. Возникшее случайно неравновесие углубилось последующей конкуренцией и отбором, в результате которого “выжили” L–аминокислоты и D–сахара.
2. Второе направление построено на концепции химической эволюции и утверждает идею некоего усложнения возникшей в ходе природного органического синтеза молекулы, одним из этапов которого и было конструирование пространственно–диссимметричной молекулы, ставшей матрицей последующего размножения подобных молекулярных систем [44].
И все же возникает вопрос: неужели столь важные вещи природа отдает на волю случая? Почему сумел накопиться один из равноценных изомеров?
Открытие несохранения четности в физике элементарных частиц показало, что природа все–таки различает правое и левое, и выбор между ними – не вполне дело случая. В середине 70–х г. ряд исследователей обратили внимание на то, что несохранение четности должно приводить к различию внутренних энергий пары оптических изомеров. В 1983 г. С.Мейзон и Дж.Трэнтер путем неэмпирического расчета простых
соединений, моделирующих структурные единицы полипептидов, показали, что L–изомер стабилизирован по сравнению с D–изомером слабыми взаимодействиями, нарушающими четность. Энергия стабилизации зависит от углов внутреннего вращения. Для углов, чаще всего встречающихся в белках, она составляет около 10–14 Дж/моль на аминокислотный остаток. Величина мизерная, однако она способна обеспечить при нормальных условиях около 106 добавочных L–макромолекул на 1 моль рацемической смеси. Пока неясно, способен ли столь слабый эффект привести к столь широкомасштабным последствиям.Живые организмы содержат большое количество киральных составных частей, но только L–аминокислоты входят в состав белков и только D–нуклеотиды находятся в нуклеиновых кислотах. Это происходит несмотря на то, что энергия обоих энантиомеров одинакова и их образование имеет равную вероятность в акиральном окружении. Тем не менее только один из них встречается в природе, и конкретные энантиомеры, характерные для жизненных процессов, одинаковы у людей,
животных, растений, микроорганизмов. Природа этого явления – одна из величайших загадок, составляющих предмет молекулярной теологии [3].Проблема происхождения биологической молекулярной стереоизомерии обрела четко выраженный междисциплинарный характер. Как сказал Д.Уолкер – редактор трудов симпозиума по происхождению молекулярной стереоизомерии, состоявшегося в июне 1979 г., – “предметом обсуждения на симпозиуме была сама междисциплинарность, поскольку простиралась она от квантовой электродинамики до химии органического синтеза... В симпозиуме приняла участие группа специалистов различных областей с целью достичь удовлетворительных знаний о том, как земная биосфера оказалась состоящей только из одного энантиомера каждой киральной молекулы” [6]. Хотя Пастер и верил, что процессы, протекающие в живой и неживой природе, разделяет пропасть, он все–таки приписывал асимметрию живой материи не “жизненной силе”, а диссимметрирующему воздействию физических факторов “космического порядка” (выражение Пастера). Согласно этой идее “дисимметрическая совокупность Вселенной” была тем самым главным физическим фактором, который предопределил зарождение биологической изомерии на молекулярном уровне [4]. В русле этих идей лежит гипотеза известного американского физика Ф.Дайсона о возникновении первых живых систем в результате своеобразного “фазового перехода” [5].
Асимметрия у животных и растений
В живой природе существуют все основные виды симметрии, которые возможны по геометрическим соображениям. То есть типы симметрии, наблюдающиеся у живых организмов, существовали в мире молекул и минералов еще до появления генов [20]. У минералов имеются оси вращательной симметрии 1–, 2–, 3–, 4– и 6–го порядков. Из этих чисел два нечетных и три четных. Симметрия 5–го порядка отсутствует
у минералов, но имеется у квазикристаллов. Она часто встречается у некоторых семейств растений, например, у Rosaceae и у иглокожих (морских ежей и морских звезд). Симметрия 3–го порядка появляется у однодольных (Iridaceae, Amarillidaceae). У цветков и беспозвоночных обычны симметрии с номером порядка, кратным порядкам у минералов: 2–, 4–, 6–, 8– и 12–го; у растений и животных появляются симметрии 3– и 6–го порядков, 5– и 10–го, а также симметрии с порядковыми числами простыми и нечетными: 7, 9, 11 и 13. То есть типы симметрии у живых организмов ничем не отличаются от тех, что существовали у минералов и квазикристаллов, они лишь “воспользовались” уже имевшимися типами симметрии или скомбинировали их, повысив порядок. По этой причине анализ симметрии в биологии ведется в разных аспектах – от оценки философского значения биологической симметрии [30] и рассмотрения ее как общебиологического явления, имеющего место на разных уровнях организации [25, 31, 32, 33], до изучения конкретных проявлений симметрии и асимметрии у различных организмов [33, 34]. В материале, который предлагается в данной статье я ограничусь рассмотрением отклонений от наиболее широко распространенной билатеральной симметрии, как симметрии левого и правого, характерной для большинства активно передвигающихся животных и ряда прикрепленных форм, обитающих на негоризонтальном субстрате [35]. При работе с биологическими объектами в настоящее время часто используется, предложенная в 1962 г. классификация симметрий–асимметрий Ван Валеном [36], согласно которой все разнообразие подразделяется на три типа:1) направленная асимметрия, когда какая–то структура развита на одной определенной стороне больше, чем на другой. В качестве примера обычно приводится сердце млекопитающих; большее развитие у одних крабов левой клешни, у других – правой; наличие лево– или правосторонней асимметрии в строении тела камбалообразных или закрученности раковины у брюхоногих моллюсков;
2) антисимметрия, характеризуемая бульшим развитием структуры то на одной, то на другой стороне тела, что соответствует отрицательной связи проявления признака на разных сторонах тела. Как пример Ван Валлен приводит левшей и правшей в популяциях человека;
3) флуктуирующая асимметрия определяется как следствие несовершенства онтогенетических процессов. По феноменологии она представляет собой незначительные ненаправленные отклонения от строгой билатеральной симметрии.
Е.Баур [21] и Г.Штуббе [22] в 30–х и 60–х гг. провели эксперименты по выяснению возможной генетической детерминации симметрии–асимметрии у биологических объектов. На одном из растений – львином зеве (
Antirrhinum) – было показано, что тип симметрии может контролироваться специальными “генами симметрии” [23]: мутация одного гена изменяла двустороннюю симметрию цветка на радиальную симметрию 5–го порядка. Причем эта мутация, затрагивающая симметрию оказывает настолько сильный эффект, что морфологические изменения достигают макроэволюционного значения: одним скачком мутант переходит в другое семейство. Такого же типа изменение симметрии наблюдается у близкого рода Linaria [23]. И все же некоторые авторы склоняются к выводу, что ген не в состоянии определять типы симметрии, а лишь осуществляет между ними выбор [20, 196]. Этот же вывод, что ген не порождает асимметрии, а только закрепляет выбор типа асимметрии, был сделан при интерпретации наследования лево– и правозакрученности раковины у пресноводного моллюска Limnaea. Раковина у этой улитки имеет форму правой спирали, но у некоторых разновидностей спираль левая. Направление спирали определяется на самых ранних стадиях эмбриогенеза и различимо уже при втором дроблении. Необычность наследования этого признака заключается в том, что он становится выраженным только во втором поколении. Так происходит по той причине, что направление закручивания определяется генами не самого индивидуума, а материнскими генами, т.е. генами ооцита, от которого особь произошла [24]. Гены воздействуют на цитоплазму яйца в яичнике, определяя направление клеточного деления, а тем самым и типа асимметрии будущего организма.Наличие асимметрии в строении тела наблюдается также у насекомых и позвоночных. Иногда асимметричность может проявляться своеобразно: встречаются насекомые, у которых сочетаются признаки самца и самки – так называемые гинандроморфы. Например, у гинандроморфного насекомого
Pseudomethoca canadensis тело как бы разделено на две половины: одна половина – крылатый самец, другая половина – бескрылая самка.Асимметрия в проявлении признаков может наблюдаться и у земноводных. Так, у некоторых видов лягушек
Dendrobates tinctorius [25] асимметрична топография распределения кожного пигмента.Некоторые экспериментальные данные
В 1980 г. в Институте цитологии и генетики СО РАН начался и продолжается по настоящее время многолетний эксперимент по селекционному преобразованию поведения у американской норки (
Mustela vison) [37]. Одним из последствий такого длительного отбора по поведению было изменение окраски у животных в ряду поколений. Изменение окраски проявлялось в неспецифическом проявлении белых отметин или пегостей на теле у животных, отбираемых на ручное поведение по отношению к человеку. Топография таких окрасочных новообразований на туловище не была хаотичной – проявление их носило направленный асимметричный характер: достоверно чаще они регистрировались на левой половине тела [45].Зарегистрированное явление побудило изучить топографию пегостей и степень их проявления у норок в промышленных популяциях, не затронутых специальным отбором на ручное или агрессивное поведение по отношению к человеку. Исследование выявило, что пегости у таких норок чаще располагаются асимметрично, т.е. площадь депигментации на одной из сторон тела достоверно превышает таковую на другой, и лишь треть животных характеризуется билатеральной симметрией в степени депигментации (рис
. 1).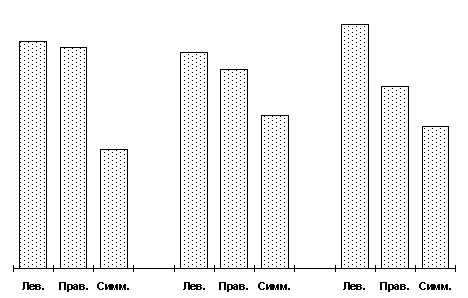
|
Кол–во |
400 |
385 |
213 |
136 |
120 |
103 |
606 |
453 |
333 |
|
% |
40 |
39 |
21 |
38 |
35 |
27 |
43 |
32 |
25 |
|
|
Селекция на |
Промышленная |
Селекция на |
||||||
Рис
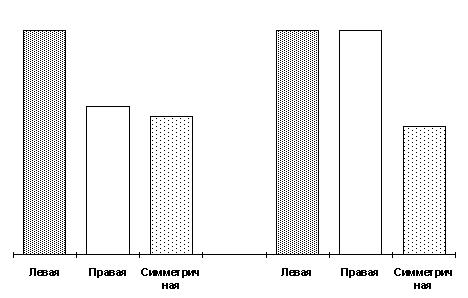
. 1. Процентное соотношение левых, правых и симметричных форм в популяциях норок,Асимметрия может быть лево– или правосторонней, а количество левых и правых форм животных примерно одинаково. Как видно из рис. 2, и у самок и у самцов наименьшая степень проявления пегостей выявлена у билатерально симметричных форм. Левосторонне асимметричные животные во всех случаях имеют максимальную площадь белой пятнистости, а вот правые формы, как видно из того же рис. 2, в степени проявления признака ведут себя двояко в зависимости от пола: у самцов они приближаются к симметричным, у самок, наоборот – к левым формам.
|
Кол–во |
71 |
68 |
68 |
65 |
52 |
35 |
|
M ± m |
4,8 ± 0,4 |
3,3 ± 0,3 |
3,1 ± 0,3 |
4,8 ± 0,5 |
4,7 ± 0,6 |
2,8 ± 0,3 |
|
s 2 |
12,9 |
5,8 |
4,5 |
16,1 |
19,2 |
7,0 |
|
|
САМЦЫ |
САМКИ |
||||
Рис
.2. Влияние пола на степень проявления признака у левых, правых и симметричных форм.Вернемся вновь к рис. 1, который показывает, что количество асимметричных форм в промышленной популяции, не затронутой специальным отбором по поведению, вдвое превышает количество симметричных. Но число норок с лево– и правосторонней асимметрией одинаково. Однако эта картина меняется в процессе отбора по поведению. В агрессивной группе уменьшается число симметричных, но соотношение животных с разносторонней асимметрией практически не меняется. Однако это соотношение изменилось в ходе отбора на доместикационное или ручное поведение: количество левых форм, как по самцам, так и по самкам достоверно превышает количество правых. То есть, в ходе направленного отбора на доместикацию, асимметрия в проявлении пегостей также приобретает направленный характер: существенно возрастает доля левых форм.
Литературные данные указывают на безуспешность попыток переделать в процессе отбора флуктуирующий характер асимметрии в направленный. На этот счет уместно привести высказывание известного эволюциониста Р.Левонтина: “В сущности, у дрозофилы нет ни одного признака – морфологического, физиологического, поведенческого или цитологического, – на который нельзя было бы вести отбор. Известна только одна неудачная попытка – при попытке Мэйнарда Смита и Сондхи отобрать признак по его левостороннему проявлению. Хотя им и удалось усилить асимметрию, все же это была флуктуирующая асимметрия без тенденции к смещению влево или вправо” [46, 47].
Трудно спекулировать на тему о том, почему проявление пегостей при отборе на доместикационное поведение носит преимущественно левосторонний характер, но в ходе этого эксперимента было зарегистрировано еще одно явление: отбор на ручное поведение повлек за собой также увеличение плодовитости и резкое возрастание изменчивости в потомстве по самым разнообразным признакам [48]. То есть количество и разнообразие вариантов, выбрасываемых в среду повышалось, и это оказалось скоррелированным с возрастанием количества левых форм. Было решено проверить, не различаются ли по плодовитости левые, правые и симметричные формы уже изначально в исходной популяции без предварительного отбора по поведению. Как показывают данные Табл. 1 левые формы норок дают преимущества в размножении.
Таблица 1. Различия в плодовитости у левых, правых и симметричных форм норок
(зверосовхоз “Знаменский” Тверской области).
|
Топография пегостей |
Количество самок |
Зарегистрировано норчат всего |
Количество норчат на 1 самку |
|
Левые |
217 |
1582 |
7.3 ± 0.3 |
|
Правые |
129 |
870 |
6.7 ± 0.2 |
|
Симметричные |
371 |
2299 |
6.2 ± 0.2 |
Складывается впечатление, что левые формы получают селективное преимущество, выбрасывая на суд естественного отбора большее количество вариантов, повышая шанс выжить в целом всей системе. Так, здесь интересно будет привести диалог между известным американским химиком Г.Вальдом и А.Эйнштейном: “Как же объяснить выбор? Почему существуют организмы с L–аминокислотами, а с D не существуют, или с D–рибофуранозами, а не с L?”
“Вы знаете, – ответил Эйнштейн – я много думал над тем, как могло случиться, что электрон отрицательный. Отрицательный, положительный – это симметрично в физике. Нет причин выбрать одно из них. И знаете к какому выводу я пришел после долгих размышлений? Я думаю – они победили в борьбе...” [50]. Г.Вальд рассуждает, что если только предположить, что киральная чистота биологических полимеров необходима организмам для нормального функционирования, то в результате борьбы за существование киральность мономеров, используемых в выживших организмах, будет стандартизирована не только в каждом отдельном биополимере, но и в организме, и в популяции организмов – во всей биосфере
. “В конкурентном мире выживает лишь тот цех, в котором стандартизирован тип используемых винтов, а организм живет в конкурентном мире” [51]. Транслировалась ли способность “выжить” с ядерного уровня на молекулярный и организменный, обеспечив тем самым преимущества левых форм, обеспечив “левый марш жизни?” На этот счет интересны наблюдения на организменном уровне среди левшей и правшей в человеческой популяции. Так, если среди недоношенных детей (вес тела до 1 кг при рождении) 54% левшей, то среди детей родившихся в срок, их уже 8 – 15% . Среди 20–летних левшей 13%, 50–летних – 5%, среди 90–летних левши уже не встречаются [52, 53]. Снова создается впечатление, что леворукость ограничивает шансы в борьбе за выживание.Селективное преимущество L–листьев фасоли над D–листьями была продемонстрирована Ю.А.Урманцевым: L–листья превосходят D–листья по интенсивности фотосинтеза и дыхания, а также по содержанию пигментов – хлорофиллов
a и b, лютеина, виолоксантина и каротина, они обладают также большей засухо– и жароустойчивостью. L–листья не только превосходят D–формы по весу и площади, но также встречаются в 2,2 раза чаще [54].В том, что L–формы в этой конкурентной борьбе могли оказаться
“временными победителями” говорит интересный факт существования фермента оксидазы D–аминокислот, найденной у организмов всех уровней организации – от бактерий до млекопитающих. Эта оксидаза катализирует окислительное дезаминирование только D–аминокислот и практически неактивна по отношению к L–аминокислотам. Несмотря на безусловное существование D–аминокислотной оксидазы, путь ее появления в природе остается тайной. Можно полагать, что наличие этого фермента свидетельство тому, что сложившееся соотношение L– и D–изомеров в современной биосфере явление не постоянное и “маятник может качнуться в противоположную сторону”.Вновь вернемся к левым и правым формам норок согласно топографии пегостей на их туловище.
В распределении пегостей по туловищу норок я рискну провести параллели с идеей, высказанной Пастером в 1850 г., когда он вводит понятие “молекулярной гемиэдрии”, с помощью которой он представляет структуру диссимметричной молекулы, проводя аналогию с гемиэдрией кристаллической. Более того, не ограничившись этой аналогией, Пастер высказался о конкретной конфигурации этой “молекулярной гемиэдрии” – “молекулярном тетраэдре” [16]. Используя тетраэдр в качестве геометрической модели оптически активных молекул, Пастер говорит: “Их, молекул, активных самих по себе можно отобразить в виде неправильного тетраэдра, инверсия которого всегда возможна” [17,
237]. То есть Пастер здесь дает указание не только на конфигурацию диссимметрической молекулы, но и на возможность инверсии тетраэдра, что соответствовало его понятию “диссимметрия”, предусматривающему отсутствие плоскости симметрии, но наличие инверсионных, то есть зеркально–поворотных осей. В 1853 г. он дает описание тетраэдной формы оптически активной диссимметричной молекулы: “Форму молекулы активного вещества можно отобразить в виде неправильных тетраэдров”. Далее, в лекции о теории молекулярной диссимметрии, прочитанной в Парижском химическом обществе 20 января 1860 г., он ставит вопрос о структуре диссимметрического расположения атомов в молекуле оптически активной винной кислоты: “Мы знаем, с одной стороны, что строение молекул двух винных кислот диссимметрично, а с другой – что они совершенно идентичны, за исключением лишь одного между ними различия, а именно, что их диссимметрии противоположны. Группируются ли атомы правой кислоты по завиткам спирали слева направо, или они находятся на вершинах неправильного тетраэдра, или расположены в соответствии с какой–либо иной диссимметрической фигурой”? И уже 3 февраля он дает ответ на поставленный вопрос: “Представьте себе винтовую лестницу, ступенями которой являются кубы. Раскрутите лестницу, и диссимметрия исчезнет. Диссимметрия лестницы – это результат способа соединения между собой этих элементарных ступеней. Таков кварц. Кристалл кварца – это вполне выстроенная лестница. Он гемиэдричен и поэтому активен в отношении поляризованного луча. Но если кристалл растворится, расплавится или его физическая структура будет разрушена то диссимметрия исчезнет, а вместе с ней и действие на поляризованный луч. Представьте, наоборот, ту же закрученную лестницу, но со ступенями в форме неправильных тетраэдров. Разрушьте лестницу, но диссимметрия сохранится, поскольку вы имеете дело с совокупностью тетраэдров [18]. С помощью такой “молекулярной механики” Пастер разработал конфигурационную модель целостной диссимметричной молекулы, в которой расположение атомов подобно тетраэдрам [19, 176].Воспользовавшись этой аналогией, я хочу предложить модель пространственного распределения пегостей по туловищу животного не хаотично, а в виде спирали с трансляцией по длине или в виде “винтовой лестницы депигментации”.
Винтовые спиральные линии иногда путают с плоскими спиралями. Спиральную форму имеют: хорошо известная раковина головоногого
Nautilus [27], колония беспозвоночного Heliodoma (Bryozoa) [27], раскрывающийся лист растения Drozophilum lusitanicum (Droseraceae) [28]. Плоские спирали проявляются в скоплениях молекул вещества, из которого образовались галактики [26]. Иногда бывает видно множество взаимопересекающихся спиралей: таковы сложные рисунки структуры цветка подсолнечника и шишки сосны строение которых было проанализировано математически [29]. Форму винтовой спирали имеют клетки водоросли Spirogyra со спиральными хлоропластами, раковина морской улитки Vermicularia spirata, рога североамериканского ископаемого быка эпохи миоцена Ilingoceros. Двойную винтовую спираль образует макромолекула ДНК; такую же, но еще более простую геометрию имеет полипептидная a–спираль молекулы кератина.Винтовое движение представляет собой соединение поворота вокруг некоторой оси с переносом вдоль этой оси. Под действием соответствующего непрерывного и равномерного движения любая точка, не лежащая на оси этого движения, описывает винтовую линию. Такое спиральное расположение часто можно наблюдать у листьев растительных побегов. Было обнаружено, что дроби вида
m/n, соответствующему винтообразному расположению листьев на стебле растения, часто являются “числами Фибоначчи”: 1/1, 1/2, 2/3, 3/5, 5/8, 8/13, 13/21, 21/34, ..., которые получаются при разложении в непрерывную дробь иррационального числа (Применимы ли “числа Фибоначчи” не только к винтообразному расположению листьев на стебельке растения, но и к распределению пегостей по туловищу норок в ходе селекционного эксперимента покажут дальнейшие исследования.
Задумываясь над фундаментальным и прикладным аспектами явления симметрии–асимметрии в попытке под “социальный заказ” выдать сиюминутную практическую отдачу научного исследования будем всегда помнить рассуждение гениального Луи Пастера, который в 1882 году уже на склоне лет говорил: “Позитивизм не подсказал мне ни одной идеи. Он грешит не только ошибками метода. Он страдает и значительными проблемами... заключающимися в том, что позитивизм не придает должного значения подлинному знанию, которое является бесконечным
” [10, 15].Литература
1.
Джаффре Г., Орчин М. Симметрия в химии. М., Мир. 1967. С. 14.2.
Гильде В. Зеркальный мир. М., Мир. 1982.3. Prelog V. Science. 1976. V. 193, P. 17.
4. Paster L. C. R.
Acad. Sci. Paris, June 1, 1874. (цит. по Haldane J.B.S., Nature, 1976. V. 185, P. 87.5. Dyson F. J. A model for the origin of Life // J. Mol. Evol. 1982. V. 18. P. 344 – 350.
6. Walker D. Origin of optical activity in nature. Preface. Amsterdam; Oxford; N.Y.: Univ. press, 1969. 257 p.
7. Pasteur Valery–Radot L. Introduction. Pasteur L. OEuvres. P.: Mason, et 1922. T. 1. P. I–XVI.
8. Dagognet Fr. Mйthodes et doctrines dans l`oeuvre de Pasteur. P.: Press univ. 1967. P. 5.
9. Морозов Л.Л. Несохраняющаяся четность в молекулярном мире организмов // Природа. 1977. № 1, С. 32 – 45.
10. Pasteur L.
Discours prononcйe dans la sйance publique tenue par ѕAcadйmie franзaise le 27 avtil 1882. P., 1882.11. Papagiannis H.D. The importance of the physical laws on the origin and evolution of life // Origins of life. Tokyo: Jap. Sci. Soc. press. 1978. P. 575 – 581.
12. Pivie N.W. Fact and assumption in studies on the origin of life // Orig. Life. 1985. V. 15. P. 207 – 212.
13. Search for the universal ancestors // Ed. H.Hartman, J.G.Lawless, P.Morrison Wash. (D. C.): NASA Sci. and Techn. Inform. Branch, 1985. 129 p.
14. Thaxton C.B., Bradley W.L., Olsen R.L. The misteryof life`s origin: Reassesing current theories. N.Y.: Phylos. Library. 1984. 317 p.
15.
Гейзенберг В. Физика и философия. Часть и целое. М., Наука. 1989. С. 342.16. Pasteur L. Recherchers sur les relations qui peuvent exister entre la forme cristalline, la composition chimique et le sens de la polarisation rotatoire. 1848 // Ibid. P. 81 – 82.
17. Pasteur L.
Nouvelles recherches sur les relations qui peuvent exister entre la forme cristalline, la composition chimique et le phйnomйne rotatoire moleculaire. 1850 // Ibid. P. 201 – 241.18. Pasteur L. Recherches sur
la dissymйtrie molйculaire des produitesorganiques naturels (Leзons professйes б la Sociйtй chimique dela Paris, le 20 janvier et le 3 fйvrier. 1860) // Ibid. P. 314 – 345.19. Pasteur L.
Mйmoir sur l`alcool amylique. 1853 // Ibid. P. 175 – 183.20. Lima–de–Faria A.
Evolution without selecton. Form and Function by Autoevolution. Institute of Molecular Cytogenetics, University of Lund, Sweden. 1988. Elsevier. Amsterdam. New York. Oxford. (А. Лима–де–Фариа. Эволюция без отбора. Автоэволюция формы и функции. Москва. Мир. 1991).21. Baur E.
Einfьhrung in die Vererbungslehre. Verlag von Gebrьder Borntraeger, 1930. Berlin.22. Stubbe H. Genetic und Zytologie von Antirrhinum L. sect. Antirrhinum. G. Fisher. 1966. Jena.
23. Gustaffson A. Linnaeus’ peloria: the history of a monster // Theor. Appl. Genet. 1979. 54. P. 241 – 248.
24. Sturtevant A.H., Beadle G.W. An Introduction to Genetics. W.B. Saunders, Philadelphia, London. 1940. P. 327 – 331.
25. Neville A.C. Animal Asymmetry. The Institute of Biology’s Stud
ies in Biology. Edward Arnold, London, 1976. № 67. P. 1 – 60.26. Burgel B.H. Los Mundos Lejanos. Labor, Barselona 1943.
27. Barnes R.D. Invertebrate Zoology. Saunders College, Philadelphia, 4th edn., 1980. P. 1 – 1089.
28. Goebel K. Organographie der Pflanzen, Dritter Teil, Samenpflancen. Gustav. Fisher, Jena. 1933.
29. Denferr D. von, Schmacher W., Magdefrau K., Ehrendorfer F. Strasburger’s Textbook of Botany. Longman, London, New York. 1971. P. 1 – 877.
30.
Урманцев Ю.А. Симметрия природы и природа симметрии. М., Мысль. 1974.31. Ludwig W. Das Rechts–Links Problem im Tierreich und beim Menchen. B.: Springer. 1932. 496 S.
32.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. М., Наука. 1964. Т. 1. 432 с.33.
Захаров В.М. Асимметрия животных. М., Наука. 1987.34. Danforth C.H. The heredity of unilateral in man // Genetics (US). 1924. V. 9
, № 3. P. 199 – 211.35.
Гиляров М.С. О функциональном значении симметрии организмов // Зоол. журн. 1944. Т. 23, вып. 5. С. 213 – 215.36. Van Valen L. A study of fluctuating asymmetry // Evolution. 1962. V. 16
, №2. P. 125 – 142.37.
Трапезов О.В. Селекционное преобразование оборонительной реакции на человека у Американской норки (Mustela vison Schreber) // Генетика. 1987. Т.23. № 6. С. 1120 – 1127.38. Puitti A.
Ьber ein neues Asparagin // Ber. Dt. chem. Ges. 1886. Bd. 19. S. 618 – 623.39.
Вейль Г. Симметрия. М., Наука, 1968.40. Enrlich F.
Ьber das Natьrlich Isomerie des Leucins // Ber. Dt. chem. Ges. 1904. Bd. 37. S. 809 – 840; 1907. Bd. 40. S. 2538 – 2562.41. Neuberg C., Mayer P.
Ьber D–, L– und R–protein cystin // Ztschr. physiol. Chem. 1905. Bd. 44. S. 498 – 510.42. Riemschneider R. Configuration et action de certains insecticides. Examen stereochimique et toxicologique d`analogue du DDT // Chim. et ind. 1954. V. 72. P. 261 – 270; 435 – 443.
43.
Болдуин Э. Основы динамической биохимии. М., Изд–во иностр. лит., 1949. 506 с.44.
Опарин А.И. Жизнь, ее природа, происхождение и развитие. М., Изд–во АН СССР, 1960. 190 с.45.
Трапезов О.В. Нарушение билатеральной симметрии в проявлении пегостей у американской норки в ходе селекционного преобразования поведения // Изв. Сиб. отд. АН СССР. 1989. Вып. 2. С. 5 – 6.46.
Левонтин Р. Генетические основы эволюции. М., Мир. 1978. 344 с.47. MaynarD–Smith J., Sondhi K.C. The genetics of a pattern // Genetics. 1960. V. 45
. № 8. P. 1039 – 1050.48.
Беляев Д.К., Трапезов О.В. Связь селекционного изменения поведения с репродуктивной функцией у американской норки // Журн. общей биол. 1986. № 4. С. 445 – 450.49.
Шубников А.В., Копцик В.А. Симметрия в науке и искусстве. М., 1972.50. Wald G. Ann. N.Y. Acad. Sci., 1957. V. 69. P. 35.
51. Pirie N.W. Trans. Bose Research Inst. 1959. V. 22. P. 11.
52. O`Callagham M.J., Tudehope D.L. // Za
ncot. 1987. № 8542. P. 1155.53. Halpern D.F., Coren S. Nature. 1988. V. 333
. № 6170. P. 213.54.
Урманцев Ю.А. Физиология растений. 1970. Т. 17. № 5. С. 937 – 944.
Институт цитологии и генетики
Сибирское отделение РАН
г. Новосибирск